Проводящая система сердца
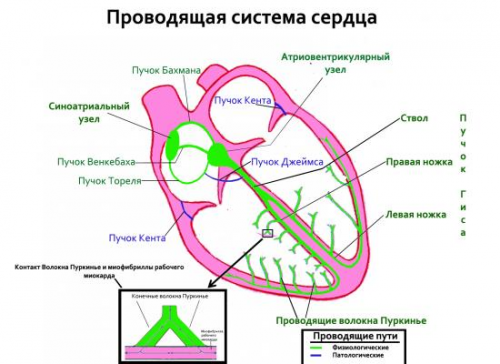
Основным координатором насосной функции предсердий и желудочков является проводящая система сердца, которая благодаря своей электрической активности способна обеспечить их согласованную работу. В норме электрический импульс генерируется в синусовом узле и активизирует оба предсердия. Наряду с этим импульс из синусового узла поступает к AV-соединению, в котором происходит некоторая задержка его продвижения, позволяющая желудочкам «без спешки» полноценно и своевременно заполниться кровью, поступающей из предсердий. Затем после прохождения AV сигнал достигает предсердно-желудочкового пучка Гиса и наконец по ветвям и волокнам Пуркинье направляется к желудочкам для активации их насосной функции.
Предсердия и желудочки разделены электрически инертными волокнистыми структурами (кольцами) так, что электрическое соединение между предсердиями и желудочками сердца при нормальных условиях обеспечивает только лишь AV-узел. Его участие в передаче сигналов позволяет предсердиям и желудочкам синхронизировать свою работу и, кроме того, минимизировать вероятность электрической обратной связи между сердечными камерами.
Проводящая система сердца представляет собой комплекс структурнофункциональных образований сердца (узлов, пучков и волокон), состоящих из атипичных мышечных волокон (син.: сердечные проводящие кардиомиоциты). Выделяют два взаимосвязанных компонента проводящей системы: синоатриальный (синусно-предсердный) и атриовентрикулярной (предсердно-желудочковый) .
Синоатриальный компонент включает синусовый узел, находящийся в стенке правого предсердия, межпредсердные пучки и межузловые проводящие тракты, связывающие предсердия друг с другом, а также с атриовентрикулярным узлом.
Синусовый узел
Синусовый узел (синузел синоатриальный, синоаурикулярный, Кисса—Флека) представлен небольшими атипичными (несократительными) кардиомиоцитами, входящими в проводящую систему сердца. Связь синусового узла с атриовентрикулярным узлом обеспечивается тремя трактами: передним (пучок Бахмана), средним (пучок Венкебаха) и задним (пучок Тореля). Обычно импульсы достигают атриовентрикулярного узла по переднему и среднему трактам. Следуя по ним, импульсы равномерно охватывают возбуждением прилегающие к проводящим путям отделы миокарда. Пейс-мекерные клетки синусового узла не имеют быстрых Na+-каналов, поэтому развивают лишь низкую скорость нарастания потенциала действия, величина которой зависит от внутриклеточного притока Са++. Вместе с тем, клетки синусового узла обладают относительно быстрой спонтанной деполяризацией (фаза 4), что обеспечивает их способность автоматически генерировать до 100 импульсов и более в минуту.
Синусовый узел богато иннервирован симпатическими и парасимпатическими нервами, которые позволяют центральной нервной системе (ЦНС) оказывать на него существенное регулирующее влияние в интересах организма.
Симпатическая стимуляция вызывает в пейсмекерных клетках повышение скорости продолжительного тока кальция. Это изменение связано с увеличением активности цАМФ и протеинкиназы А, которое обусловливает фосфорилирование Ca++-L каналов. Симпатическая стимуляция увеличивает также ток калия из клетки, что укорачивает продолжительность потенциала действия и способствует преждевременному старту следующего потенциала действия.
Наконец, симпатическая стимуляция увеличивает вход Na+ в клетку, что приводит к повышению скорости спонтанной диастолической деполяризации. Активация парасимпатической нервной системы вызывает противоположный эффект. Увеличение ацетилхолина активирует G-белок, который ингибирует аденилатциклазу и приводит к снижению концентрации цАМФ, что уменьшает скорость ионных потоков кальция в клетку, калия из клетки и натрия в клетку.
Предсердно-желудочковый компонент объединяет расположенный в нижней стенке правого предсердия атриовентрикулярный узел и отходящий от него пучок Гиса, который имеет 2 ножки — правую и левую. Этот пучок связывает между собой желудочки. Отходящие от пучка Гиса ветви обозначают как волокна Пуркинье.
В атриовентрикулярном АВ-соединении, главным образом в его пограничных участках между атриовентрикулярным узлом и пучком 1иса, происходит достаточно существенное замедление скорости проведения импульсов. Эта замедление обеспечивает отсроченное возбуждение желудочков после окончания полноценного сокращения предсердий. В целом основными функциями атриовентрикулярного узла являются:
а) антеградная задержка и «фильтрация» волн возбуждения от предсердий к желудочкам, обеспечивающая скоординированное сокращение предсердий и желудочков;
б) функциональная защита желудочков от возбуждения в «уязвимой» фазе потенциала действия: минимизация вероятности электрической обратной связи между желудочками и предсердиями.
Кроме того, в условиях угнетения активности синоатриального узла атриовентрикулярный узел способен выполнять роль самостоятельного генератора сердечного ритма, т.е. выступать в качестве пейсмекера второго порядка, индуцируя в среднем 40—60 импульсов в минуту.
Доминирующим в роли пейсмекера при прочих равных условиях является синусовый узел – водитель ритма первого порядка, т.к. в норме по сравнению с АВ-узлом генерирует импульсы с большей частотой.
Атриовентрикулярный узел
Атриовентрикулярный (АВ) узел (син.: АВ узел Ашоффа—Тавары; АВ-соединение). Предсердия изолированы от желудочков фиброзным кольцом, которое неспособно пропускать сигналы от синусового узла. В норме есть только один электрически активный путь между предсердиями и желудочками — это атриовентрикулярный узел, нередко называемый АВ-соединением В предсердной части АВ-узла находятся т. н. «переходные» клетки-пейсмекеры, аналогичные клеткам водителя ритма первого порядка. Скорость (крутизна) спонтанной диастолической деполяризации в этих клетках очень низкая, составляя всего 0,05 м/с (для сравнения скорость проведения сигналов в предсердии 1,0 м/с), поэтому пороговый потенциал возбуждения достигается медленнее, что можно объяснить, во-первых, исключительно продолжительным током кальция в клетки-пейсмекеры, а во-вторых, — их низкой плотностью в АВ-соединении.
Пучок Гиса (син.: АВ-пучок Гиса) и волокна Пуркинье (син.: система Шса-Пуркинье). Пучок Гкса — это совокупность волокон, которые заключены в фиброзные оболочки и отходят от АВ-узла, постепенно расслаиваясь на две группы волокон—левую ножку пучка, которая иннервирует межжелудочковую перегородку, левый желудочек, и правый пучок, иннервирующий правый желудочек. Дистальные ветви этих пучков проникают во все регионы правого и левого желудочков, образуя систему Пуркинье.
Потенциалы действия пучка Шса и волокон Пуркинье схожи между собой. Для них характерны быстрая фаза 0 деполяризации, длительный период плато, и очень медленная диастолическая деполяризация. Быстрая фаза 0 деполяризации обусловлена чрезвычайно высокой плотностью быстрых Na+-каналов. Длительный период плато (фаза 2), как полагают, возникает либо из-за сравнительно поздней инактивации Са2+-каналов или поздней активации К+-каналов. Фаза 4 деполяризации замедлена из-за медленного потока ионов Na+ внутрь клетки (If). Достаточно быстрое проведение сигналов в системе Пуркинье необходимо для практически одновременной активации желудочков. Этому способствует также высокая плотность синаптических контактов клеток Пуркинье на кардиомиоцитах (рис. 6.9).
Проводящая система обладает рядом свойств, определяющих ее участие в работе сердца: автоматизм, возбудимость и проводимость. Основным из них является автоматизм, без которого остальные свойства бессмысленны.
Автоматизм клеток миокарда
Автоматизм — это способность специализированных клеток миокарда спонтанно вырабатывать электрические импульсы (син: потенциалы действия; ПД). Существует продольный (от предсердий к верхушке сердца) градиент автомата и проводящей системы. Принято различать три «центра» автоматизма:
1. синоатриальный узел — водитель ритма сердца первого порядка. В физиологических условиях этот узел генерирует импульсы с частотой 60-1 80 в мин;
2. атриовентрикулярный узел (клетки АВ-соединения) – водитель ритма сердца второго порядка, который способен генерировать 40—50 импульсов в 1 мин;
3. пучок Гиса (30—40 импульсов в 1 мин) и волокна Пуркинье (в среднем 20 импульсов в 1 мин) — водители ритма третьего порядка.
В норме единственным водителем ритма является синоатриальный узел, 1 который «не позволяет» реализоваться автоматической активности других потенциальных водителей ритма.
В основе автоматизма лежит медленная диастолическая деполяризация, постепенно понижающая мембранный потенциал до уровня порогового (критического) потенциала, с которого начинается быстрая регенеративная деполяризация мембраны, или фаза 0 потенциала действия.
Ритмичное возбуждение пейсмекерных клеток с частотой 70—80 в 1 мин можно объяснить двумя процессами: 1) ритмичным спонтанным повышением проницаемости мембран этих клеток для ионов Na+ и Са++, вследствие чего они поступают в клетку; 2) ритмичным снижением проницаемости для J ионов К+, в результате чего количество покидающих клетку ионов К+ уменьшается.
Согласно предложенному недавно альтернативному механизму, входящий пейсмекерный ток ионов Na+ (If) со временем возрастает, тогда как выходящий ток К+ остается неизменным. В целом данные процессы детерминируют развитие мед ленной диастолической деполяризации клеток пейс-мекера и достижение критического порога возбуждения (—40 мВ), обеспечивающего возникновение потенциала действия и его распространение по миокарду. Восходящая часть ПД клеток-пейсмекеров обеспечивается входом Са2+ в клетку Отсутствие плато можно объяснить характерным изменением проницаемости мембраны для ионов, при котором процессы деполяризации и инверсии плавно переходят в реполяризацию, которая также проходит более медленно из-за замедленного тока К+ из клетки. Амплитуда ПД составляет 70—80 мВ, его продолжительность — около 200 мс, рефрактерность — около 300 мс, те. длительность рефрактерного периода продолжительнее ПД, что защищает сердце от внеочередных импульсов (и соответственно преждевременного возбуждения), исходящих из других (как нормальных, так , и патологических) генераторов возбуждения, приходящихся на период не-возбудимости сердечной мышцы.
Функционирование дистальной (эффекторной) часта проводящей системы обеспечивают такие же процессы, которые происходят в клетках сино-атриального пейсмекера. В развитии спонтанной диастолической депаляризации в структурах системы Гиса—Пуркинье важную роль играет также ток ионов Na+ (И). Кроме того, в этом процессе участвуют и другие ионные токи, включая ток ионов К+ (ik), который в значительной степени определяет зависимость автоматизма волокон Пуркинье от внеклеточной концентрации ионов К+. При этом, отметим ток ионов К+ весьма незначителен в пейсмекерных клетках синоатриального узла, поскольку в них мало калиевых каналов.
В современной модели автоматизма волокон Пуркинье представлены четыре ионных механизма, зависящие от внеклеточной концентрации ионов К+:
1) активация тока ионов Na+ (If), усиливающая пейсмекерную активность;
2) активация тока ионов К+ (Ik), замедляющая или приостанавливающая пейсмекерную активность;
3) активация Na+/K+-Hacoca (Ip), замедляющая пейсмекерную активность;
4) уменьшение тока ионов K+(Ik), усиливающая пейсмекерную активность.
С электрофизиологической точки зрения, интервал между сокращениями сердца равен отрезку времени, в течение которого мембранный потенциал покоя в клетках-пейсмекерах синоатриального узла смещается до уровня порогового потенциала возбуждения
Существует строгая согласованность между процессом электрической активации каждого кардиомиоцита [потенциалом действия], возбуждением всего миокардиального синцития [ЭКГ-комплексом] и сердечным циклом [биомеханограммой] сердца.
 (No Ratings Yet)
(No Ratings Yet)
